Haplogroup J1 – PPNB/Northeast Caucasians
Haplogroup J2 – Hurrians/Northeast Caucasians – Kura Araxes
Haplogroup G – Northwest Caucasians – Maykop
Haplogroup R1b
-
Haplogroup R1b
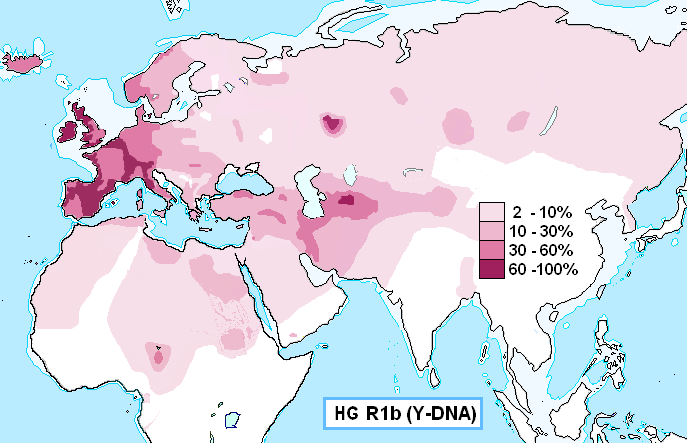
While the Hurrians are connected with Haplogroup J2, the Indo-Europeans is connected with Haplogroup R1b, the most common haplogroup in Western Europe, reaching over 80% of the population in Ireland, the Scottish Highlands, the Western Wales, the Atlantic fringe of France, the Basque country and Catalonia. It is also common in Anatolia and around the Caucasus, in parts of Russia and in Central and South Asia.
Haplogroup R* originated in North Asia just before the Last Glacial Maximum (26,500-19,000 years ago). This haplogroup has been identified in the remains of a 24,000 year-old boy from the Altai region, in south-central Siberia. This individual belonged to a tribe of mammoth hunters that may have roamed across Siberia and parts of Europe during the Paleolithic.
Autosomally this Paleolithic population appears to have contributed mostly to the ancestry of modern Europeans and South Asians, the two regions where haplogroup R also happens to be the most common nowadays (R1b in Western Europe, R1a in Eastern Europe, Central and South Asia, and R2 in South Asia).
The oldest forms of R1b (M343, P25, L389) are found dispersed at very low frequencies from Western Europe to India, a vast region where could have roamed the nomadic R1b hunter-gatherers during the Ice Age. The three main branches of R1b1 (R1b1a, R1b1b, R1b1c) all seem to have stemmed from the Middle East.
The southern branch, R1b1c (V88), is found mostly in the Levant and Africa. The northern branch, R1b1a (P297), seems to have originated around the Caucasus, eastern Anatolia or northern Mesopotamia, then to have crossed over the Caucasus, from where they would have invaded Europe and Central Asia. R1b1b (M335) has only been found in Anatolia.
Domestication of cattle
It has been hypothetised that R1b people (perhaps alongside neighbouring J2 tribes) were the first to domesticate cattle in northern Mesopotamia some 10,500 years ago. Cattle herders probably maintained a nomadic or semi-nomadic existence, while other people in the Fertile Crescent (presumably represented by haplogroups E1b1b, G and T) settled down to cultivate the land or keep smaller domesticates.
The analysis of bovine DNA has revealed that all the taurine cattle (Bos Taurus) alive today descend from a population of only 80 aurochs. The earliest evidence of cattle domestication dates from circa 8,500 BCE in the Pre-Pottery Neolithic cultures in the Taurus Mountains.
Pre-Pottery Neolithic B (PPNB) is a division of the Neolithic developed by Kathleen Kenyon during her archaeological excavations at Jericho in the southern Levant region.
Cultural tendencies of this period differ from that of the earlier Pre-Pottery Neolithic A (PPNA) period in that people living during this period began to depend more heavily upon domesticated animals to supplement their earlier mixed agrarian and hunter-gatherer diet.
Sites from this period found in the Levant utilizing rectangular floor plans and plastered floor techniques were found at Ain Ghazal, Yiftahel (western Galilee), and Abu Hureyra (Upper Euphrates). The period is dated to between ca. 10,700 and ca. 8,000 BP or 7000 – 6000 BCE.
Tell Aswad, a large agricultural village between Mount Hermon and Damascus, have a fully established PPNB culture at 8700 BC, pushing back the period’s generally accepted start date by 1,200 years.
Similar sites to Tell Aswad in the Damascus Basin of the same age were found at Tell Ramad and Tell Ghoraifé. How a PPNB culture could spring up in this location, practicing domesticated farming from 8700 BC has been the subject of speculation.
Like the earlier PPNA people, the PPNB culture developed from the Earlier Natufian but shows evidence of a northerly origin, possibly indicating an influx from the region of north eastern Anatolia.
The culture disappeared during the 8.2 kiloyear event, a term that climatologists have adopted for a sudden decrease in global temperatures that occurred approximately 8,200 years before the present, or c. 6200 BCE, and which lasted for the next two to four centuries.
In the following Munhatta and Yarmukian post-pottery Neolithic cultures that succeeded it, rapid cultural development continues, although PPNB culture continued in the Amuq valley, where it influenced the later development of Ghassulian culture.
Work at the site of ‘Ain Ghazal in Jordan has indicated a later Pre-Pottery Neolithic C period which existed between 8,200 and 7,900 BP. Juris Zarins has proposed that a Circum Arabian Nomadic Pastoral Complex developed in the period from the climatic crisis of 6200 BCE, partly as a result of an increasing emphasis in PPNB cultures upon animal domesticates, and a fusion with Harifian hunter gatherers in Southern Palestine, with affiliate connections with the cultures of Fayyum and the Eastern Desert of Egypt. Cultures practicing this lifestyle spread down the Red Sea shoreline and moved east from Syria into southern Iraq.
The two oldest archaeological sites showing signs of cattle domestication are the villages of Çayönü Tepesi, a Neolithic settlement in southeastern Turkey inhabited around 7200 to 6600 BC., and Dja’de el-Mughara in northern Iraq, two sites only 250 km away from each other. This is presumably the area from which R1b lineages started expanding – or in other words the “original homeland” of R1b.
Haplogroup J2 is thought to have appeared somewhere in the Middle East towards the end of the last glaciation, between 15,000 and 22,000 years ago. Its present geographic distribution argues in favor of a Neolithic expansion from the Fertile Crescent.
This expansion probably correlated with the diffusion of domesticated of cattle and goats (starting c. 8000-9000 BCE) from the Zagros Mountains and northern Mesopotamia, rather than with the development of cereal agriculture in the Levant (which appears to be linked rather to haplogroups G2 and E1b1b).
A second expansion of J2 could have occured with the advent of metallurgy, notably copper working (from the Lower Danube valley, central Anatolia and northern Mesopotamia), and the rise of some of the oldest civilisations.
Quite a few ancient Mediterranean and Middle Eastern civilisations flourished in territories where J2 lineages were preponderant. This is the case of the Hattians, the Hurrians, the Etruscans, the Minoans, the Greeks, the Phoenicians (and their Carthaginian offshoot), the Israelites, and to a lower extent also the Romans, the Assyrians and the Persians. All the great seafaring civilisations from the middle Bronze Age to the Iron Age were dominated by J2 men.
There is a distinct association of ancient J2 civilisations with bull worship. The oldest evidence of a cult of the bull can be traced back to Neolithic central Anatolia, notably at the sites of Çatal Höyük and Alaca Höyük. Bull depictions are omnipresent in Minoan frescos and ceramics in Crete. Bull-masked terracotta figurines and bull-horned stone altars have been found in Cyprus (dating back as far as the Neolithic, the first presumed expansion of J2 from West Asia). The Hattians, Sumerians, Babylonians, Canaaites, and Carthaginians all had bull deities (in contrast with Indo-European or East Asian religions).
The sacred bull of Hinduism, Nandi, present in all temples dedicated to Shiva or Parvati, does not have an Indo-European origin, but can be traced back to Indus Valley civilisation. Minoan Crete, Hittite Anatolia, the Levant, Bactria and the Indus Valley also shared a tradition of bull leaping, the ritual of dodging the charge of a bull. It survives today in the traditional bullfighting of Andalusia in Spain and Provence in France, two regions with a high percentage of J2 lineages.
Notwithstanding its strong presence in West Asia today, haplogroup J2 does not seem to have been one of the principal lineages associated with the rise and diffusion of cereal farming from the Fertile Crescent and Anatolia to Europe. The region of origin of J2 is still unclear at present. It is likely that J2 men had settled over most of Anatolia, the South Caucasus and Iran by the end of the Last Glaciation 12,000 years ago.
It is possible that J2 hunter-gatherers then goat/sheep herders also lived in the Fertile Crescent during the Neolithic period, although the development of early cereal agriculture is thought to have been conducted by men belonging primarily to haplogroups E1b1b and G2a.
The first expansion of J2 into Europe probably happened during the Late Glacial and immediate postglacial periods (16.000-10.000), when Anatolian hunter-gatherers moved into the Balkans. This migration would have included J2b and E-V13 male lineages and assorted J and T maternal lineages (presumably J1c, J2a1, T1a1, T2a1b, T2b, T2e and T2f1).
This population would have occupied the modern regions of western Turkey, Greece, Macedonia, Albania, Serbia and Bulgaria. When farmers and herders expanded from the Fertile Crescent during the Early Neolithic, they would have blended with one another before expanding towards Central Europe and Italy.
J2b seems to have split in two subclades soon after leaving Anatolia, with J2b1 being found mostly in western Anatolia and Greece, while J2b2 expanded from the Balkans to most of Europe, to Central Asia, India and back to the Middle East.
It is very likely that J2a, J1 and G2a were the three dominant male lineages the Early Bronze Age Kura-Araxes culture, which expanded from the South Caucasus to eastern Anatolia, northern Mesopotamia and the western Iran. From then on, J2 men would have definitely have represented a sizeable portion of the population of Bronze and Iron Age civilizations such as the Hurrians, the Assyrians or the Hittites.
Within India, J2a is more common among the upper castes and decreases in frequency with the caste level. This can be explained by the assimilation of local J2a (and R2) people from Bactria and Pakistan by the R1a Indo-European warriors who descended from the Volga-Ural region of Russia (Sintashta culture) and established themselves for a few centuries in southern Central Asia, immediately north of the Hindu Kush (including the Oxus civilization) before moving on to conquer the Indian subcontinent.
J2a would have reached Bactria with the expansion of Neolithic herders from the Middle East who then blended with the indigenous hunter-gatherers belonging chiefly to R2.
J2b has a quite different distribution from J2a. J2b seems to have a stronger association with the Neolithic and Chalcolithic cultures of Southeast Europe. It is particularly common in the Balkans, Central Europe and Italy, which is roughly the extent of the European Copper Age culture. Its maximum frequency is achieved around Albania, Kosovo, Montenegro and Northwest Greece – the part of the Balkans which best resisted the Slavic invasions in the Early Middle Ages.
The vast majority of J2b lineages belong to J2b2 and its subclades. While J2b* and J2b1 lineages are mostly restricted to the Caucasus, Anatolia and the Balkans, J2b2 is also found in the Pontic Steppe, in Central Asia and in South Asia, particularly in India. It’s very low frequency in the Middle East though suggests that unlike other J2 lineages it was not disseminated by a demic diffusion of the Neolithic lifestyle.
In many ways the distribution of J2b2 and its subclades is strongly reminiscent of G2a3b1 and its subclades. The most likely hypothesis is that both haplogroups penetrated into the Pontic Steppe region during the Neolithic or Chalcolithic period, either crossing the Caucasus from eastern Anatolia or, more probably, expanding east from the flourishing cultures of ‘Old Europe’, either as an early expansion of the Thessalian Neolithic or as part of the Carpatho-Balkan Metallurgical Province. J2b2 and G2a3b1 would have integrated the local R1a and R1b tribes in the Pontic steppe and the North Caucasus.
Nowadays J2b2 is found chiefly in south-east and Central Europe, but also in Russia and throughout the Indian subcontinent. All these elements reinforce the hypothesis that J2b2 and G2a3b1 were two minor lineages spread within an R1a-dominant population during the Indo-Aryan invasions of South Asia approximately 3,500 years ago.
Another conceivable possibility is that a minority of J2b2, G2a3b1 and R1b-M269 from the Caucasus region migrated to the Volga-Ural region in the early Bronze Age, propagating with them the Proto-Indo-European language and bronze technology to the Caspian steppe before the expansion of this new culture to Central and South Asia. The drawback of this hypothesis is that it doesn’t explain why R1b lineages strongly outnumber J2b2 and G2a3b1 in Europe but not in South Asia.
The Western European R1b is dominated by specific sub-clades of R-M269, with some small amounts of other types found in areas such as Sardinia. Within Europe, R-M269 is dominated by R-M412, also known as R-L51, which is virtually absent in the Near East, the Caucasus and West Asia. This Western European population is further divided between R-P312/S116 and R-U106/S21, which appear to spread from the western and eastern Rhine river basin respectively.
As of early 2014, there were 286 mutations (SNPs) defining haplogroup G, confirming that this paternal lineage experienced a severe bottleneck before splitting into happlogroups G1 and G2. Haplogroup G1 might have originated around modern Iran, while G2 would have developed in Southwest Asia during the Upper Paleolithic, probably in the Late Glacial period (19,000 to 12,000 years ago). At that time humans would all have been hunter-gatherers and in most cases been living in small nomadic or semi-nomadic tribes.
Members of haplogroup G2 appear to have been closely linked to the development of early agriculture in the Levant part of the Fertile Crescent, starting 11,500 years before present. The G2a branch expanded to Anatolia, the Caucasus and Europe, while G2b ended up secluded in the southern Levant and is now found mostly among Jewish people.
There has so far been ancient Y-DNA analysis from only four Neolithic cultures (LBK in Germany, Remedello in Italy and Cardium Pottery in south-west France and Spain), and all sites yielded G2a individuals, which is the strongest evidence at present that farming originated with and was disseminated by members of haplogroup G (although probably in collaboration with other haplogroups such as E1b1b, J, R1b and T).
The highest genetic diversity within haplogroup G is found between the Levant and the Caucasus, in the Fertile Crescent, which is a good indicator of its region of origin. It is thought that early Neolithic farmers expanded from the Levant and Mesopotamia westwards to Anatolia and Europe, eastwards to South Asia, and southwards to the Arabian peninsula and North and East Africa.
The domestication of goats and cows first took place in the mountainous region of eastern Anatolia, including the Caucasus and Zagros. This is probably where the roots of haplogroup G2a (and perhaps of all haplogroup G) are to be found. So far, the only G2a people negative for subclades downstream of P15 or L149.1 were found exclusively in the South Caucasus region.
The early R1b cattle herders would have split in at least three groups. One branch (M335) remained in Anatolia, but judging from its extreme rarity today wasn’t very successful, perhaps due to the heavy competition with other Neolithic populations in Anatolia, or to the scarcity of pastures in this mountainous environment.
A second branch migrated south to the Levant, where it became the V88 branch. Some of them searched for new lands south in Africa, first in Egypt, then colonizing most of northern Africa, from the Mediterranean coast to the Sahel.
The third branch (P297), crossed the Caucasus into the vast Pontic-Caspian Steppe, which provided ideal grazing grounds for cattle. They split into two factions: R1b1a1 (M73), which went east along the Caspian Sea to Central Asia, and R1b1a2 (M269), which at first remained in the North Caucasus and the Pontic Steppe between the Dnieper and the Volga.
It is not yet clear whether M73 actually migrated across the Caucasus and reached Central Asia via Kazakhstan, or if it went south through Iran and Turkmenistan. In the latter case, M73 might not be an Indo-European branch of R1b, just like V88 and M335.
R1b-M269 (the most common form in Europe) is closely associated with the diffusion of Indo-European languages, as attested by its presence in all regions of the world where Indo-European languages were spoken in ancient times, from the Atlantic coast of Europe to the Indian subcontinent, which comprised almost all Europe (except Finland, Sardinia and Bosnia-Herzegovina), Anatolia, Armenia, European Russia, southern Siberia, many pockets around Central Asia (notably in Xinjiang, Turkmenistan, Tajikistan and Afghanistan), without forgetting Iran, Pakistan, northern India and Nepal. The history of R1b and R1a are intricately connected to each other.
Kur
The Neolithic, Eneolithic and early Bronze Age cultures in Pontic-Caspian steppe has been called the Kurgan culture (4200-2200 BCE) by Marija Gimbutas, due to the lasting practice of burying the deads under mounds (“kurgan”) among the succession of cultures in that region.
In Sumerian mythology, Kur is considered the first ever dragon, and usually referred to the Zagros Mountains to the east of Sumer. The cuneiform for “kur” was written ideographically with the cuneiform sign 𒆳, a pictograph of a mountain. It can also mean “foreign land”.
Although the word for earth was Ki, Kur came to also mean land, and Sumer itself, was called “Kur-gal” or “Great Land”. “Kur-gal” also means “Great Mountain” and is a metonym for both Nippur and Enlil who rules from that city. Ekur, “mountain house” was the temple of Enlil at Nippur. A second, popular meaning of Kur was “underworld”, or the world under the earth.
Kur was sometimes the home of the deads, it is possible that the flames on escaping gas plumes in parts of the Zagros mountains would have given those mountains a meaning not entirely consistent with the primary meaning of mountains and an abode of a god. The eastern mountains as an abode of the gods are popular in Ancient Near Eastern mythology.
The underworld Kur is the void space between the primeval sea (Abzu) and the earth (Ma). Kur is almost identical with “Ki-gal”, “Great Land” which is the Underworld (thus the ruler of the Underworld is Ereshkigal “Goddess of The Great Land”. In later Babylonian myth Kur is possibly an Anunnaki, brother of Ereshkigal, Inanna, Enki, and Enlil.
In the Enuma Elish in Akkadian tablets from the first millennium BC, Kur is part of the retinue of Tiamat, and seems to be a snakelike dragon. In one story the slaying of the great serpent Kur results in the flooding of the earth.
A first millennium BC cylinder seal shows a fire-spitting winged dragon—a nude woman between its wings—pulling the chariot of the god who subdued it, another depicts a god riding a dragon, a third a goddess.
It is likely that this name coincides with the modern day Kurds who are the predominant ethnicity inhabiting much of the Zagros mountain range.Hennerbichler believes the term Kurd and similar ethnic labels to have been derived from the Sumerian word stem “kur”, meaning mountain. In modern Kurdish language, kur is the word for boy.
Kurgan culture
Kurgan is the Turkic term for a tumulus, derived from the Iranic Scythian root, gur+khan[a] (“grave house”, mausoleum). In Latvian Kurm – means “mound, hill” which corresponds to Sumerian KUR.MA “a star rising”, all with the same basic root kur – “hill, mound” as in Kurgan.
It is now known that kurgan-type burials only date from the 4th millenium BCE and almost certainly originated south of the Caucasus. The genetic diversity of R1b being greater around eastern Anatolia, it is hard to deny that R1b evolved there before entering the steppe world.
Kurgans were built in the Eneolithic, Bronze, Iron, Antiquity and the Middle Age, with old traditions still active in Southern Siberia and Central Asia. A plethora of place names that include the word “kurgan” appear from Lake Baikal to the Black Sea, and includes the city Kurgan and the administrative center of Kurgan Oblast located in Southern Russia as part of the Urals Federal District.
Burial mounds are complex structures with internal chambers. Within the burial chamber at the heart of the kurgan, elite individuals were buried with grave goods and sacrificial offerings, sometimes including horses and chariots. Kurgans were used in the Ukrainian and Russian Steppes but spread into eastern, central, and northern Europe in the 3rd millennium BC.
In Kurgan cultures, most of the burials were in kurgans, either clan kurgans or individual ones. Most prominent leaders were buried in individual kurgans, now called “Royal kurgans”, which attract the greatest attention and publicity. Kurgan barrows were characteristic of Bronze Age peoples, from the Altay Mountains to the Caucasus, Ukraine, Romania, and Bulgaria.
Depending on a combination of elements, each historical and cultural nomadic zone has its architectural peculiarities. The structures of the earlier Neolithic period, from the 4th to the 3rd millenniums BC., and Bronze Age until the 1st millennium BC., display continuity of the archaic forming methods driven by the common ritual-mythological ideas.
The Kurgan hypothesis
The Neolithic, Eneolithic and early Bronze Age cultures in Pontic-Caspian steppe has been called the Kurgan culture (7000-2200 BCE) by Marija Gimbutas, due to the lasting practice of burying the deads under mounds (“kurgan”) among the succession of cultures in that region.
Marija Gimbutas writes that the Kurgans (Yamnaya or Pit-Grave culture) ostensibly started their migrations in Southern Russia and the Ukrainian steppes at the Black Sea, reaching Anatolia and Mesopotamia through the Balkans and Greece, across the Caucasus then east to Central Asia and south into Iran. These migrations would account for all the mound-grave cultures we know, even those surfacing in China (through the Tocharians) and in the Indus Valley at around this time.
The Kurgan hypothesis postulates that the Proto-Indo-Europeans were the bearers of the Kurgan culture of the Black Sea and the Caucasus and west of the Urals. The hypothesis was introduced by Marija Gimbutas in 1956, combining kurgan archaeology with linguistics to locate the origins of the Proto-Indo-European (PIE) speaking peoples. She tentatively named the culture “Kurgan” after their distinctive burial mounds and traced its diffusion into Europe.
Those scholars who follow Gimbutas identify a “Kurgan culture” as reflecting an early Indo-European ethnicity which existed in the steppes and southeastern Europe from the 5th to 3rd millennia BC.
Marija Gimbutas’ Kurgan hypothesis is opposed by Paleolithic Continuity Theory, which associates Pit Grave and Sredny Stog Kurgan cultures with Turkic peoples, and the Anatolian hypothesis, and is also opposed by the Black Sea deluge theory.
The distribution of such tumuli in Eastern Europe corresponds closely to the area of the Pit Grave or Kurgan culture in South-Eastern Europe. Kurgan cultures are divided archeologically into different sub-cultures, such as Timber Grave, Pit Grave, Scythian, Sarmatian, Hunnish and Kuman-Kipchak.
Modern linguists have placed the Proto-Indo-European homeland in the Pontic-Caspian steppe, a distinct geographic and archeological region extending from the Danube estuary to the Ural Mountains to the east and North Caucasus to the south.
The Kura–Araxes culture
The Kura–Araxes culture or the Early Trans-Caucasian culture was a civilization that existed from 3400 BC until about 2000 BC, which has traditionally been regarded as the date of its end, but it may have disappeared as early as 2600 or 2700 BC.
The earliest evidence for this culture is found on the Ararat plain; thence it spread to Georgia by 3000 BC (but never reaching Colchis), and during the next millennium it proceeded westward to the Erzurum plain, southwest to Cilicia, and to the southeast into an area below the Urmia basin and Lake Van, and finally down to the borders of present day Syria. Altogether, the early Trans-Caucasian culture, at its greatest spread, enveloped a vast area approximately 1,000 km by 500 km.
The name of the culture is derived from the Kura and Araxes river valleys. Its territory corresponds to parts of modern Armenia, Georgia, Azerbaijan, Chechnya, Dagestan, Ingushetia and North Ossetia. It may have given rise to the later Khirbet Kerak ware culture found in Syria and Canaan after the fall of the Akkadian Empire.
Archaeological evidence of inhabitants of the Kura–Araxes culture had shown that ancient settlements were found along the Hrazdan river, as shown by drawings at a mountainous area in a cave nearby.
Structures within settlements have not revealed much differentiation, nor was there much difference in size or character between settlements, facts that suggest they probably had a poorly developed social hierarchy for at least a significant stretch of their history. Some, but not all, settlements were surrounded by stone walls.
They built mud-brick houses, originally round, but later developing into subrectangular designs with structures of just one or two rooms, multiple rooms centered around an open space, or rectilinear designs.
At some point the culture’s settlements and burial grounds expanded out of lowland river valleys and into highland areas. Although some scholars have suggested that this expansion demonstrates a switch from agriculture to pastoralism, and that it serves as possible proof of a large-scale arrival of Indo-Europeans, facts such as that settlement in the lowlands remained more or less continuous suggest merely that the people of this culture were diversifying their economy to encompass both crop and livestock agriculture.
The economy was based on farming and livestock-raising (especially of cattle and sheep). They grew grain and various orchard crops, and are known to have used implements to make flour. They raised cattle, sheep, goats, dogs, and in its later phases, horses (introduced around 3000 BC, probably by Indo-European speaking tribes from the North.
There is evidence of trade with Mesopotamia, as well as Asia Minor. It is, however, considered above all to be indigenous to the Caucasus, and its major variants characterized (according to Caucasus historian Amjad Jaimoukha) later major cultures in the region.
The extent of the Kuro-Araxes culture (light shading) shown in relation to subsequent cultures in the area, such as Urartu (dark shading).
In its earliest phase, metal was scant, but it would later display “a precocious metallurgical development which strongly influenced surrounding regions”. They worked copper, arsenic, silver, gold, tin, and bronze.
Their metal goods were widely distributed, recorded in the Volga, Dnieper and Don-Donets systems in the north, into Syria and Palestine in the south, and west into Anatolia.
Their pottery was distinctive; in fact, the spread of their pottery along trade routes into surrounding cultures was much more impressive than any of their achievements domestically. It was painted black and red, using geometric designs for ornamentation. Examples have been found as far south as Syria and Israel, and as far north as Dagestan and Chechnya.
The spread of this pottery, along with archaeological evidence of invasions, suggests that the Kura-Araxes people may have spread outward from their original homes, and most certainly, had extensive trade contacts.
Jaimoukha believes that its southern expanse is attributable primarily to Mitanni and the Hurrians.
They are also remarkable for the production of wheeled vehicles (wagons and carts), which were sometimes included in burial kurgans. Inhumation practices are mixed. Flat graves are found, but so are substantial kurgan burials, the latter of which may be surrounded by cromlechs.
Late in the history of this culture, its people built kurgans of greatly varying sizes, containing greatly varying amounts and types of metalwork, with larger, wealthier kurgans surrounded by smaller kurgans containing less wealth.
This trend suggests the eventual emergence of a marked social hierarchy. Their practice of storing relatively great wealth in burial kurgans was probably a cultural influence from the more ancient civilizations of the Fertile Crescent to the south.
This points to a heterogeneous ethno-linguistic population. Hurrian and Urartian elements are quite probable, as are Northeast Caucasian ones. Some authors subsume Hurrians and Urartians under Northeast Caucasian as well as part of the Alarodian theory.
The presence of Kartvelian languages was also highly probable. Influences of Semitic languages and Indo-European languages are also highly possible, though the presence of the languages on the lands of the Kura–Araxes culture is more controversial. In the Armenian hypothesis of Indo-European origins, this culture (and perhaps that of the Maykop culture) is identified with the speakers of the Anatolian languages.
The Maykop culture
The Maykop (also spelled Maikop) culture (3700-2500 BC), in the North Caucasus, was a major Bronze Age archaeological culture in the Western Caucasus region of Southern Russia and was culturally speaking a sort of southern extension of the Yamna horizon. Radiocarbon dates for various monuments of the Maykop culture are from 3950 – 3650 – 3610 – 2980 calBC.
It extends along the area from the Taman Peninsula at the Kerch Strait to near the modern border of Dagestan and southwards to the Kura River. The culture takes its name from a royal burial found in Maykop in the Kuban River valley.
In the south it borders the approximately contemporaneous Kura-Araxes culture (3500—2200 BC), which extends into eastern Anatolia and apparently influenced it. To the north is the Yamna culture, including the Novotitorovka culture (3300—2700), which it overlaps in territorial extent. It is contemporaneous with the late Uruk period in Mesopotamia.
The Maykop kurgan was extremely rich in gold and silver artifacts; unusual for the time. Its inhumation practices were characteristically Indo-European, typically in a pit, sometimes stone-lined, topped with a kurgan (or tumulus). Stone cairns replace kurgans in later interments.
In the early 20th century, researchers established the existence of a local Maykop animal style in the found artifacts. This style was seen as the prototype for animal styles of later archaeological cultures: the Maykop animal style is more than a thousand years older than the Scythian, Sarmatian and Celtic animal styles. Attributed to the Maykop culture are petroglyphs which have yet to be deciphered.
The Maykop culture is believed to be one of the first use the wheel. The Maykop nobility enjoyed horse riding and probably used horses in warfare. It should be noted that the Maykop people lived sedentary lives, and horses formed a very low percentage of their livestock, which mostly consisted of pigs and cattle.
Archaeologists have discovered a unique form of bronze cheek-pieces, which consists of a bronze rod with a twisted loop in the middle and a thread through her nodes that connects with bridle, halter strap and headband. Notches and bumps on the edges of the cheek-pieces were, apparently, to fix nose and under-lip belts.
The construction of artificial terrace complexes in the mountains is evidence of their sedentary living, high population density, and high levels of agricultural and technical skills. The terraces were built around the fourth millennium BC., and all subsequent cultures used them for agricultural purposes.
The vast majority of pottery found on the terasses are from the Maykop period, the rest from the Scythian and Alan period. The Maykop terraces are among the most ancient in the world, but they are little studied. The longevity of the terraces (more than 5000 years) allows us to consider their builders unsurpassed engineers and craftsmen.
The Kura Araxes culture is closely linked to the approximately contemporaneous Maykop culture of Transcaucasia. As Amjad Jaimoukha puts it: “The Kura-Araxes culture was contiguous and had mutual influences with the Maikop culture in the Northwest Caucasus”.
According to E.I.Krupnov (1969:77), there were elements of the Maikop culture in the early memorials of Chechnya and Ingushetia in the Meken and Bamut kurgans and in Lugovoe in Serzhen-Yurt.
Similarities between some features and objects of the Maikop and Kura-Araxes cultures, such as large square graves, the bold-relief curvilinear ornamentation of pottery, ochre-colored ceramics, earthen hearth props with horn projections, flint arrowheads, stone axes and copper pitchforks are indicative of a cultural unity that pervaded the Caucasus in the Neolithic Age.»
The Kuban River is navigable for much of its length and provides an easy water-passage via the Sea of Azov to the territory of the Yamna culture, along the Don and Donets River systems. The Maykop culture was thus well-situated to exploit the trading possibilities with the central Ukraine area.
New data revealed the similarity of artifacts from the Maykop culture with those found by the ancient city of Tell Khazneh in northern Syria, the construction of which dates back to 4000 BC.
In 2010, nearly 200 Bronze Age sites were reported stretching over 60 miles between the Kuban and Nalchik rivers, at an altitude of between 4,620 feet and 7,920 feet. They were all “visibly constructed according to the same architectural plan, with an oval courtyard in the center, and connected by roads.”
“The work unearthed the remains of hundreds of tombs and ancient settlements from the end of the Copper Age (Chalcolithic or Eneolithic Age, the first half of the 4th millennium BC) right up to the Middle Ages inclusive. These sites were found mainly in Western Azerbaijan, along the middle reaches of the Kur River, from 4350 until 4000 BC. From ancient times the favorable geography of the region attracted tribes engaged in agriculture and cattle-breeding.
These finds are evidence of the high development of crafts such as pottery, bone working, stone working, metal working and weaving of the settled agricultural and cattle-breeding tribes. The Soyugbulag barrows, the first barrows of the Copper Age in the whole South Caucasus, are especially important remains of the Leyla Tepe culture.
Apart from Khojakhan, all the aforementioned Copper Age sites concern the Leyla Tepe archaeological culture or people. Therefore, the research into the remains of the Leyla Tepe culture is of great importance in the study of the cultural, economic and ethnic links of the Caucasus and the Middle East.
The Leyla-Tepe culture includes a settlement in the lower layer of the settlements Poilu I, Poilu II, Boyuk-Kesik I and Boyuk-Kesik II. They apparently buried their dead in ceramic vessels. Similar amphora burials in the South Caucasus are found in the Western Georgian Jar-Burial Culture.
The culture has also been linked to the north Ubaid period monuments, in particular, with the settlements in the Eastern Anatolia Region (Arslan-tepe, Coruchu-tepe, Tepechik, etc.). The settlement is of a typical Western-Asian variety, with the dwellings packed closely together and made of mud bricks with smoke outlets
After inhabiting the Northern Caucasus in the second half of the 4th millennium BC, the Leyla Tepe people became part of the Maykop people. It has been suggested that the Leyla-Tepe were the founders of the Maykop culture.
An expedition to Syria by the Russian Academy of Sciences revealed the similarity of the Maykop and Leyla-Tepe artifacts with those found recently while excavating the ancient city of Tel Khazneh I, from the 4th millennium BC.
The Indo-Europeans
Because of its burial practices the Maykop culture is in terms of the Kurgan hypothesis of Marija Gimbutas seen as an Indo-European intrusion from the Pontic steppe into the Caucasus. However, as J. P. Mallory point out that: «… where the evidence for barrows is found, it is precisely in regions which later demonstrate the presence of non-Indo-European populations.»
The culture has been described as, at the very least, a “kurganized” local culture with strong ethnic and linguistic links to the descendants of the Proto-Indo-Europeans. It has been linked to the Lower Mikhaylovka group and Kemi Oba culture, and more distantly, to the Globular Amphora and Corded Ware cultures, if only in an economic sense.
However, according to J.P.Mallory: «Such a theory, it must be emphasized, is highly speculative and controversial although there is a recognition that this culture may be a product of at least two traditions: the local steppe tradition embraced in the Novosvobodna culture and foreign elements from south of the Caucasus which can be charted through imports in both regions.»
Gamkrelidze and Ivanov, whose views are somewhat controversial, suggest that the Maykop culture (or its ancestor) may have been a way-station for Indo-Europeans migrating from the South Caucasus and/or eastern Anatolia to a secondary Urheimat on the steppe.
This would essentially place the Anatolian stock in Anatolia from the beginning, and at least in this instance, agrees with Colin Renfrew’s Anatolian hypothesis. Considering that some attempt has been made to unite Indo-European with the Northwest Caucasian languages, an earlier Caucasian pre-Urheimat is not out of the question. However, most linguists and archaeologists consider this hypothesis highly unlikely, and prefer the Eurasian steppes as the genuine IE Urheimat.
Although not generally considered part of the Pontic-Caspian steppe culture due to its geography, the North Caucasus had close links with the steppe, as attested by numerous ceramics, gold, copper and bronze weapons and jewelry in the contemporaneous cultures of Mikhaylovka, Sredny Stog and Kemi Oba.
The link between the North Pontic and North Caucasus is older than the Maykop period. Its predecessor, the Svobodnoe culture (4400-3700 BC), already had links to the Suvorovo-Novodanilovka and early Sredny Stog cultures, and the even older Nalchik settlement (5000-4500 BC) displayed a similar culture as Khvalynsk on the Volga. This may be the period when R1b started interacting and blending with the R1a population of the steppes.
The Yamna and Maykop people both used kurgan burials, with their deads in a supine position with raised knees and oriented in a north-east/south-west axis. Graves were sparkled with red ochre on the floor, and sacrificed dometic animal buried alongside humans.
They also had in common horse riding, wagons, a cattle- and sheep-based economy, the use of copper/bronze battle-axes (both hammer-axes and sleeved axes) and tanged daggers. In fact, the oldest wagons and bronze artefacts are found in the North Caucasus, and spread from there to the steppes.
Maykop was an advanced Bronze Age culture, actually one of the very first to develop metalworking, and therefore metal weapons. The world’s oldest sword was found at a late Maykop grave in Klady kurgan 31. Its style is reminiscent of the long Celtic swords, though less elaborated.
Horse bones and depictions of horses already appear in early Maykop graves, suggesting that the Maykop culture might have been founded by steppe people or by people who had close link with them.
However, the presence of cultural elements radically different from the steppe culture in some sites could mean that Maykop had a hybrid population. Without DNA testing it is impossible to say if these two populations were an Anatolian R1b group and a G2a Caucasian group, or whether R1a people had settled there.
The two or three etnicities might even have cohabited side by side in different settlements. Typical Caucasian Y-DNA lineages (such as G2a) do not follow the pattern of Indo-European migrations, so intermarriages must have been limited, or at least restricted to Indo-European men taking Caucasian wives rather than the other way round.
Maykop people are the ones credited for the introduction of primitive wheeled vehicles (wagons) from Mesopotamia to the steppes. This would revolutionize the way of life in the steppe, and would later lead to the development of (horse-drawn) war chariots around 2000 BC.
Cavalry and chariots played an vital role in the subsequent Indo-European migrations, allowing them to move quickly and defeat easily anybody they encountered. Combined with advanced bronze weapons and their sea-based culture, the western branch (R1b) of the Indo-Europeans from the Black Sea shores are excellent candidates for being the mysterious Sea Peoples, who raided the eastern shores of the Mediterranean during the second millennium BC.
Modern linguists have placed the Proto-Indo-European homeland in the Pontic-Caspian Steppe, a distinct geographic and archeological region extending from the Danube estuary to the Ural Mountains to the east and North Caucasus to the south.
Horses were first domesticated around 4000 BC in the steppe, perhaps somewhere around the Don or the lower Volga, and soon became a defining element of steppe culture.
During the Bronze-age period, known as the Yamna horizon (3300-2500 BC), the cattle and sheep herders adopted wagons to transport their food and tents, which allowed them to move deeper into the steppe, giving rise to a new mobile lifestyle that would eventually lead to the great Indo-European migrations.
The Pontic-Caspian steppe cultures can be divided in a western group, ranging from the Don River to the Dniester (and later Danube), and an eastern one, in the Volga-Ural region. The Pontic steppe was probably inhabited by men of mixed R1a and R1b lineages, with higher densities of R1b just north of the Caucasus, and more R1a in the northern steppes and the forest-steppes.
R1b almost certainly crossed over from northern Anatolia to the Pontic-Caspian steppe. It is not clear whether this happened before, during or after the Neolithic. A regular flow of R1b across the Caucasus cannot be excluded either.
The genetic diversity of R1b being greater around the Caucasus, it is hard to deny that R1b settled and evolved there before entering the steppe world.
Does that mean that Indo-European languages originated in the steppes with R1a people, and that R1b immigrants blended into the established culture? Or that Proro-Indo-European language appear in northern Anatolia or in the Caucasus, then spread to the steppes with R1b? Or else did Proro-Indo-European first appear in the steppe as a hybrid language of Caucasian/Anatolian R1b and steppe R1a?
This question has no obvious answer, but based on the antiquity and archaic character of the Anatolian branch (Hittite, Palaic, Luwian, Lydian, and so on) a northern Anatolian origin of Proto-Indo-European is credible.
Furthermore, there is documented evidence of loan words from Caucasian languages in Indo-European languages. This is much more likely to have happened if Proto-Indo-European developed near the Caucasus than in the distant steppes. Haplogroup R1b would consequently have been the spreading factor of PIE to the steppes, and from there to Europe, Central Asia and South Asia.
The maternal lineages (mtDNA) corresponding to haplogroup R1b
Haplogroup R1b is very widespread in most of Europe and across vast swathes of North Africa, the Middle East and Central Asia today. As R1b Indo-Europeans advanced from the Pontic-Caspian Steppe by marrying local women as well as Indo-European women, it is difficult to estimate what were the original mtDNA haplogroups of R1b people back in the steppes, or prior to that in the Neolithic Near East or Paleolithic Eurasia.
There are two methods to determine the haplogroups of the original Indo-Europeans. The first one is to look at ancient DNA. Hundreds of mtDNA samples from Neolithic, Mesolithic and even Paleolithic Europe have been tested to date. Some haplogroups have never been identified outside Northeast Europe before the Bronze Age. These include H2a1, H4, H6, H8, H15, I, T1, U2 and W. All of them except H8 and H15 were found in Bronze Age cultures associated with the diffusion of the Indo-Europeans speakers like Corded Ware and Unetice.
Interestingly, Mesolithic and Paleolithic Northeast Europeans were only found to belong to haplogroups U2, U4 and U5, as well as some undetermined subclades of H and T. U4 and U5 were also found in the rest of Europe before the Neolithic. So it could be that mtDNA I, T1 and W came with R1b from the Near East, but this hasn’t been confirmed yet.
It is unclear whether there was any haplogroup V before the Bronze Age either. None could be identified with certainty, but that may simply be because the defining mutations of haplogroup V lie within the coding region of mitochondrial DNA, which is more difficult to recover from ancient samples. Actually not a single V sample has shown up even in Bronze and Iron Age samples. MtDNA V being spread very evenly across all Europe nowadays, it is hard to believe that it wasn’t found among Mesolithic Europeans, and probably Indo-Europeans too.
The problem of the first method is that it only works to identify haplogroups that were brought to Europe by the Indo-Europeans during the Bronze Age, but it doesn’t take into account the haplogroups that the Indo-Europeans had in common with Mesolithic or Neolithic Europeans. For example, it is certain that the Bronze Age Indo-Europeans shared haplogroups U4 and U5 with Mesolithic hunter-gathers, and it is very likely that many H subclades were also present among Mesolithic Europeans, including H1, H3, H10 and H11.
The second method attempts to remedy to this issue by looking at the European mtDNA found in Bronze Age and modern samples in parts of Asia that were settled by the Indo-Europeans. Isolated Indo-European settlements in Asia, such as the Tarim basin in north-west China, the Altai region in southern Siberia, or Bactria and Margiana in southern Central Asia, provide unique opportunities to find European mtDNA in regions that are otherwise genetically Asian.
Since there wasn’t any other major European settlement of these regions historically, if we exclude white Russians from the sampled populations, the European mtDNA found in these regions would necessarily correlate with Bronze and Iron Age Indo-Europeans, i.e. haplogroups R1a and R1b.
The only interference could come from Middle Eastern mtDNA, especially in Muslim parts of Central Asia and in Xinjiang. Fortunately we have an idea of what mt-haplogroups could have been brought by E1b1b, J1, J2 and T1 people, such as haplogroups HV, N1, J, K, T2 and U3. The Scythians in particular are thought to have hybridized extensively with the descendants of Neolithic southern Central Asians, as ancient Scythian mtDNA contain a lot of these haplogroups (alongside H2a1, U2, U5 and East Asian lineages). But all Scythian Y-DNA so far turned out to be R1a, although they surely included R1b and J2 lineages too.
The European or Middle Eastern lineages found in Central and North Asia include HV, H2a1, H5a, H6, H7, I1, I2, I4, J1b1, J1c (incl. J1c2m), K, N1a, R, T1, T2, U1, U2e, U3, U4, U5, U8, V, W, and X2. If we exclude the haplogroups that are found mostly in R1a countries, or more typical of the Middle East than Europe, what is left as the potential maternal lineages of Bronze Age R1b men are H2a1, H5a, H6, H7, H8, I, J1, K, T1, T2, U4, U5 and V. U3, W and X2 might also be included, although they correlate less with the modern distribution of R1b. The K subclades of likely Indo-European origin include K1a3, K1a4, K1c1 and K2b.
The Indo-European tribes that carried R1b Y-chromosomes were accompanied by other populations assimilated in the Pontic Steppe, or before that in the Caucasus and eastern Anatolia. These minorities included Y-haplogroups G2a3b1, J2b2, R1a-L664 and T1a-L131 and T1a-P77. It is likely that H4, H6, U2 and U4 came from the indigenous steppe population (paternally R1a).
Haplogroups H2a1, I1, I2, I4, K1a4 and K1c1, R (mostly R1a), T2 and W is all common in the North Caucasus and could have been assimilated to the R1b population during the Maykop period. MtDNA haplogroups H5a, H7, J1c, K, T2 and X2 were all present during the Neolithic in Europe and the Near East. They could have entered the R1b gene pool in the eastern Anatolia or northern Mesopotamia during the Neolithic.
R1b tribes are thought to have domesticated cattle in that region 10,500 years ago, yet only moved across the Caucasus sometime between 7,500 and 6,500 years ago. For three or four millennia, semi-nomadic R1b herders were bound to have intermingled with some of the Near Eastern or Caucasian neighbours.
Another possibility is that these lineages came from Balkan-Carpathian farmers or herders belonging to Y-haplogroups G2a, J2b and T1a who moved to the Pontic Steppe during the Late Neolithic or Chalcolithic and who were absorbed by R1b tribes in the steppes, before the Indo-European migrations took place.
This leaves only mtDNA haplogroups H8c, H15, J1b1a, U5 and V as the most likely potential original maternal lineages of R1b tribes before they started mixing with other Near Eastern populations. Looking at deeper subclades, J1b1a and T1a1a display extremely strong correlations with the distribution of Y-haplogroup R1b.
One way of determining what mtDNA haplogroups R1b tribes carried at the very beginning of the Neolithic, 10,000 years ago, is to compare the above haplogroups with those of African ethnic groups known to possess elevated percentages of R1b-V88. The best studied group are the Fulani, whose mtDNA include three European-looking haplogroups J1b1a, U5 and V making up about 15% of their total maternal lineages.
These haplogroups have been identified in all four Central African countries sampled, confirming a strong correlation with haplogroup R1b. Since African R1b-V88 and Eurasian haplogroup R1b-P297 split roughly 10,000 years ago, there is little doubt that J1b, U5 and V were three of the original maternal lineages of R1b people. Only the J1b1b subclade seems to be related to the propagation of Y-haplogroup R1b.
Other J1b subclades are geographically restricted to the Near East, particularly from the Caucasus to the Arabian peninsula. J1b might have been the first indigenous Near Eastern lineage assimilated by R1b tribes when they moved into the region (presumably from Russia or Iran, or the Caspian Sea, which was only formed by the melting of Russian glaciers just before the Neolithic began). If that is the case, the Paleolithic R1b people would have belonged exclusively to mtDNA U5 and V, just like the modern Sami, and like a lot of Mesolithic Europeans.
R1 populations spread genes for light skin, blond hair and red hair
There is now strong evidence that both R1a and R1b people contributed to the diffusion of the A111T mutation of the SLC24A5, which explains apporximately 35% of skin tone difference between Europeans and Africans, and most variations within South Asia. The distribution pattern of the A111T allele (rs1426654) of matches almost perfectly the spread of Indo-European R1a and R1b lineages around Europe, the Middle East, Central Asia and South Asia.
The mutation was probably passed on in the Early neolithic to other Near Eastern populations, which explains why Neolithic farmers in Europe already carried the A111T allele, although at lower frequency than modern Europeans and southern Central Asians.
The light skin allele is also found at a range of 15 to 30% in in various ethnic groups in northern sub-Saharan Africa, mostly in the Sahel and savannah zones inhabited by tribes of R1b-V88 cattle herders like the Fulani and the Hausa. This would presuppose that the A111T allele was already present among all R1b people before the Pre-Pottery Neolithic split between V88 and P297. R1a populations have an equally high incidence of this allele as R1b populations.
On the other hand, the A111T mutation was absent from the 24,000-year-old R* sample from Siberia, and is absent from most modern R2 populations in Southeast India and Southeast Asia. Consequently, it can be safely assumed that the mutation arose among the R1* lineage during the late Upper Paleolithic, probably some time between 20,000 and 13,000 years ago.
Fair hair was another physical trait associated with the Indo-Europeans. In contrast, the genes for blue eyes were already present among Mesolithic Europeans belonging to Y-haplogroup I. The genes for blond hair are more strongly correlated with the distribution of haplogroup R1a, but those for red hair have not been found in Europe before the Bronze Age, and appear to have been spread primarily by R1b people.
The lactase persistence allele and R1b cattle pastoralists
Lactose (milk sugar) is an essential component of breast milk consumed by infants. Its digrestion is made possible by an enzyme, called lactase, which breaks down lactose in simple sugars that can be absorbed through the intestinal walls and into the bloodstream.
In most mammals (humans included), the production of the lactase enzyme is dramatically reduced soon after weaning. As a result, older children and adults become lactose intolerant. That is true of a big part of the world population. Some people possess a genetic mutation that allows the production of lactase through adulthood. This is called lactase persistence (LP).
Lactase persistence is particularly common among Northwest Europeans, descended from the ancient Celtic and Germanic people, and in parts of Africa where cattle herding has been practiced for thousands of years.
The highest incidence for the lactase persistence alleles, known to geneticists as -13,910*T (rs4988235) and -22018*A (rs182549), are found among Scandinavian, Dutch, British, Irish and Basque people. Sub-Saharan populations with lactase persistence have different mutations, such as -14010*C, -13915*G and -13907*G.
There has been speculations among geneticists and evolutionary biologists regarding the origin of the lactase persistence allele in Europeans. Over 100 ancient DNA samples have been tested from Mesolithic, Neolithic and Bronze Age Europe and Syria, and the -13910*T allele has been found only in Late Neolithic/Chalcolithic and Bronze Age individuals.
A paper published in 2007 by Burger et al. indicated that the genetic variant that causes lactose persistence in most Europeans (-13,910*T) was rare or absent in early farmers from central Europe. A study published by Yuval Itan and colleagues in 2010 clearly shows this. The origin of the mutation does not really matter, since it could have been present at low frequencies in the human gene pool for tens of thousands of years before it underwent postive selective pressure among cattle-herding societies.
A study published in 2009, also by Itan et al., suggests that the Linear Pottery culture (also known as Linearbandkeramik or LBK), which preceded the Funnel (-neck-) Beaker culture, short TRB or TBK from (German) Trichter (-rand-) becherkultur (ca 4300 BC–ca 2800 BC), by some 1,500 years, was the culture in which this trait started to co-evolve with the culture of dairying.
TRB was an archaeological culture in north-central Europe. It developed as a technological merger of local Neolithic and Mesolithic techno-complexes between the lower Elbe and middle Vistula rivers, introducing farming and husbandry as a major source of food to the pottery-using hunter-gatherers north of this line. It has been suggested that this culture was the origin of the gene allowing adults of Northern European descent to digest lactose.
It was claimed that in the area formerly inhabited by this culture, prevalence of the gene is virtually universal. Ancient DNA extracted from three individuals ascribed to a TRB horizon in Gökhem, Sweden, was found to possess mtDNA haplogroups H, J, and T.
In the context of the Kurgan hypothesis, the culture is seen as non-Indo-European, representing the culture of what Marija Gimbutas termed Old Europe, the peoples of which were later to be governed by the Indo-European-language-speaking peoples intruding from the east. The political relation between the aboriginal and intrusive cultures resulted in quick and smooth cultural morphosis into Corded Ware culture.
Heterodoxically, Dutch publications mention mixed burials and propose a quick and smooth internal change to Corded Ware within two generations occurring about 2900 BC in Dutch and Danish TRB territory, probably precluded by economic, cultural and religious changes in East Germany, and call the migrationist view of steppe intrusions introducing Indo-European languages obsolete (at least in this part of the world).
R1b men are thought to be the first people on earth to successfully domesticate cattle and to develop a lifestyle based on cattle husbandry and herding during the Pre-Pottery Neolithic. Looking for pasture for their cows, R1b tribes migrated from the Near East to the savannah of North Africa (which has since underwent desertification and become the Sahara) and to the Pontic Steppe in southern Russia and Ukraine. For several millennia no other human population was so depended on cattle for their survival as these R1b tribes.
It is known that most Neolithic herding societies consumed at least some animal milk and even made cheese from it (since cheese contains less lactose and is easier to digest for people who are lactose intolerant). In most of Europe, the Middle East and South Asia, people essentially herded goats and sheep, better suited to mountainous environment of the Mediterranean basin, Anatolia and Iran. Goats and sheep could also be kept easily inside villages by sedentary cereal cultivators, while cows needed vast pastures for grazing, which were particularly scare in the Middle East.
Domesticated cattle were sometimes found in small number among other Neolithic populations, but the ones that relied almost entirely on them were the R1b tribes of the Pontic Steppe and North Africa. To this very day, semi-nomadic pastoralists in the Sahel, such as the Fulani and the Hausa, who are descended from Neolithic R1b-V88 migrants from the Near East, still maintain primarily herds of cattle. It is among these cattle herders that selective pressure for lactase persistence would have been the strongest.
What is certain is that individuals from Bronze Age cultures associated with the arrival of Indo-European speakers from the Pontic Steppe already possessed relatively high percentages of the LP allele. For example the LP allele was found at a frequency of 27% among the 13 individuals from the Lichtenstein Cave in Germany, who belonged to the Urnfield culture, and were a mix of Y-haplogroups R1b, R1a and I2a2b.
Nowadays, the LP allele is roughly proportional to the percentage of R1b, and to a lower extent R1a, found in a population. In the British Isles, the Low Countries and south-west Scandinavia, where LP is the highest in the world, the combined percentage of R1a and R1b exceeds 70% of the population.
In Iberia, the highest percentage of LP is observed among the Basques, who have the highest percentage of R1b. In Italy, LP is most common in the north, like R1b. The lowest incidence of LP in Europe is found in South Italy, Greece and the Balkans, the regions that have the least R1b lineages.
Filed under: Uncategorized

